
Study on the molecular mechanism of miR-508-3p targeted regulation of paired box gene 2 affecting the biological characteristics of lung cancer cells
-
摘要:目的 探讨微小RNA-508-3p(miR-508-3p)在肺癌组织中的表达及其对肺癌细胞增殖活力、侵袭能力、凋亡率的影响与机制。方法 采用定量聚合酶链反应(qPCR)法和蛋白质印迹法(Western blotting)检测肺癌组织中miR-508-3p和配对盒基因2(PAX2)表达,分析两者的相关性。分别用miR-508-3p模拟物(miR-508-3p组)、模拟物阴性对照miR-NC(miR-NC组)、PAX2 siRNA si-PAX2 (si-PAX2组)、siRNA阴性对照(si-NC组)、miR-508-3p抑制剂anti-miR-508-3p(anti-miR-508-3p组)、抑制剂阴性对照anti-miR-NC(anti-miR-NC组)、共转染miR-508-3p和PAX2过表达质粒pcDNA-PAX2 (miR-508-3p+pcDNA- PAX2组)、共转染miR-508-3p和空白质粒pcDNA-NC(miR-508-3p+pcDNA-NC组)转染肺癌A549细胞,并设置空白组。采用噻唑蓝(MTT)法检测24~72 h的细胞增殖活力; 采用Transwell小室及流式细胞术分别检测细胞侵袭能力及凋亡率,采用Western blotting检测A549细胞基质金属蛋白酶-2(MMP-2)、B淋巴细胞瘤-2(Bcl-2)和Bcl-2相关X蛋白(Bax)蛋白表达。根据双荧光素酶活性检测结果评估miR-508-3p对PAX2的靶向调控。结果 肺癌组织miR-508-3p表达低于癌旁组织, PAX2 mRNA和PAX2蛋白表达高于癌旁组织,差异有统计学意义(P < 0.05); PAX2蛋白和miR-508-3p的表达呈显著负相关(P < 0.05)。与空白组比较, miR-508-3p组Bax表达和细胞凋亡率均升高, MMP-2、Bcl-2蛋白表达和增殖活力、侵袭能力均降低,差异有统计学意义(P < 0.05); 与si-NC组比较, si- PAX2组PAX2、MMP-2、Bcl-2表达降低, Bax表达、凋亡率升高,增殖活力和侵袭能力降低,差异有统计学意义(P < 0.05)。miR-508-3p靶向PAX2基因,调控PAX2蛋白的表达。PAX2可以逆转miR-508-3p对A549细胞增殖活力、侵袭能力以及细胞凋亡的影响。结论 miR-508-3p在肺癌组织低表达,其过表达可降低A549细胞增殖活力及侵袭能力,诱导细胞凋亡,机制与靶向调控PAX2基因有关。Abstract:Objective To investigate the expression of miR-508-3p in lung cancer tissues, explore the effects of proliferation activity, invasion ability and apoptosis rate and mechanisms.Methods Quantitative polymerase chain reaction (qPCR) and western blotting were used to detect the expressions of miR-508-3p and paired box gene 2(PAX2) in lung cancer tissues, and the correlation between them was analyzed. miR-508-3p mimics (miR-508-3p group), mimic negative control miR-NC (miR-NC group), PAX2 siRNA si- PAX2 (si- PAX2 group), siRNA negative control (si-NC group), miR-508-3p inhibitor anti-miR-508-3p (anti-miR-508-3p group), inhibitor negative control anti-miR-NC (anti-miR-NC group), co-transfected miR-508-3p and PAX2 overexpression plasmid pcDNA- PAX2 (miR-508-3p+pcDNA- PAX2 group), co-transfected miR-508-3p and blank plasmid pcDNA-NC (miR-508-3p+pcDNA-NC group) were transfected into lung cancer A549 cells and blank (NC) group was established. Methyl tetrazolium (MTT) assay was used to detect cell proliferation activity in 24 to 72 h; transwell chamber and flow cytometry were used to detect cell invasion ability and apoptosis rate, western blotting was used to detect A549 cell invasion-related matrix metalloproteinase-2 (MMP-2) and apoptosis-related B lymphocytoma-2(Bcl-2) and Bcl-2 associated X protein(Bax). The dual luciferase activity was used to evaluate the actions of targeted regulation of PAX2 by miR-508-3p.Results The expression of miR-508-3p in lung cancer tissue was lower than that in adjacent tissues, and the expression of PAX2 mRNA and PAX2 protein was significantly higher than that in adjacent tissues (P < 0.05). The expression of PAX2 protein and miR-508-3p was negatively correlated (P < 0.05). Compared with the blank group, the expression of Bax and apoptosis rate of the miR-508-3p group were increased, and the expressions of MMP-2 and Bcl-2 proteins, proliferation activity and invasion ability were decreased (P < 0.05). Compared with si-NC group, the expressions of PAX2, MMP-2 and Bcl-2 in the si- PAX2 group were decreased, the expression of Bax and apoptosis rate were increased, and the proliferation activity and invasion ability were decreased (P < 0.05). MiR-508-3p targeted PAX2 and regulated the expression of PAX2. PAX2 could reverse the effects of miR-508-3p on the proliferation, invasion and apoptosis of A549 cells.Conclusion MiR-508-3p is under-expressed in lung cancer tissues, its overexpression can reduce the proliferation and invasion of A549 cells and induce cell apoptosis. The mechanism is related to the targeted regulation of PAX2 gene.
-
Keywords:
- lung cancer /
- miR-508-3p /
- PAX2 gene /
- RNA interference /
- invasion /
- apoptosis
-
肺癌是临床最常见的恶性肿瘤之一,主要治疗途径为外科手术、放疗、化疗及生物靶向治疗,但治疗后易复发或转移,患者5年生存率低、病死率高(主要原因为肿瘤的持续增殖及转移)[1-2]。因此,研究肺癌的分子机制并寻找新的治疗靶点具有重要意义。微小RNA(miRNA)是高度保守的小型非编码RNA,长度为20~25个核苷酸,其可通过与mRNA的3′非翻译区(3′UTR)互补结合而沉默靶基因的表达,导致mRNA被降解或翻译被抑制,与癌症的发生发展有关[3]。微小RNA-508-3p(miR-508-3p)在胃癌[4]、卵巢癌[5]中表达下调并发挥肿瘤抑制功能,但其在肺癌中的作用尚不明确。配对盒基因2(PAX2)是PAX转录因子家族成员之一,在食管癌、前列腺癌等多种肿瘤中表达升高,可促进肿瘤细胞生长,参与肿瘤的发生发展[6]。研究[7]报道, PAX2在肺癌中表达升高,且淋巴结转移组PAX2表达高于非淋巴结转移组,提示PAX2可能影响肺癌细胞生长。小干扰RNA(siRNA)是近年来研究基因沉默的重要工具,可特异性抑制相应靶基因mRNA表达,进而降解靶基因,也称为转录后调控,目前已被广泛应用于肿瘤治疗、基因功能研究等多方面[8-9]。本研究检测肺癌组织miR-508-3p、PAX2表达水平,探讨miR-508-3p对肺癌细胞增殖活力、侵袭能力、凋亡的影响及对PAX2的调控作用与机制,现报告如下。
1. 材料与方法
1.1 标本及细胞
收集2018年6月—2019年3月本院胸外科手术切除的肺癌组织标本,同时收集相应的癌旁组织(距离肿瘤边缘1~3 cm)为对照,标本共来自43例患者,其中男20例、女23例,年龄36~70岁,平均58.4岁。所有患者经病理学检查确认病理类型为腺癌,且均为首次治疗,术前未接受放疗、化疗及其他抗癌治疗。所有组织标本经手术切除后置于液氮罐中保存备用。本研究已获得患者知情同意,并经医院伦理委员会审核批准。人肺腺癌A549细胞购自美国模式培养物集存库(ATCC)。
1.2 主要试剂和仪器
胎牛血清(FBS)及RPMI1640培养基均购自美国Gibco公司; PAX2、基质金属蛋白酶-2(MMP-2)、B淋巴细胞瘤-2(Bcl-2)和Bcl-2相关X蛋白(Bax)抗体均购自美国Abcam公司; 二喹啉甲酸(BCA)试剂盒购自中国碧云天公司; 聚偏氟乙烯(PVDF)膜购自上海生工生物工程有限公司; 噻唑蓝(MTT)、二甲基亚砜(DMSO)购自美国Sigma公司; 蛋白质印迹法(Western blotting)细胞裂解液购自美国Promega公司; Transwell小室购自美国corning costar公司; 细胞凋亡试剂盒及流式细胞仪均购自美国BD公司; 酶标仪购自美国Biotek公司; CO2细胞培养箱购自美国Thermo公司。
1.3 细胞培养及转染
取出保存在液氮罐中的A549细胞,迅速解冻后在CO2体积分数为5%、37 ℃细胞培养箱中,用含10%FBS的RPMI1640培养基培养细胞。取对数生长期细胞,转染前1 d, 接种细胞于24孔细胞培养板,接种密度为3×104个/mL, 用含10% FBS的RPMI1640培养基培养,次日,观察到细胞贴壁,且生长密度达60%~70%时,使用Opti-MEM培养基分别稀释si-PAX2 (转染PAX2特异性siRNA,si-PAX2组)、si-NC(转染阴性对照siRNA, si-NC组)和转染试剂,设置加脂质体的为空白组, 5 min内将siRNA与转染试剂混合,室温静置20 min, 将混合液加入培养板相应孔内,将细胞置于培养箱内常规培养, 6~8 h后,将转染液弃去,加含10% FBS的RPMI1640培养基培养。具体操作步骤参照美国Invitrogen生产的Lipofectamie 2 000转染试剂的操作说明。同法转染miR-508-3p(转染miR-508-3p模拟物, miR-508-3p组)、miR-NC(转染模拟物阴性对照miR-NC,miR-NC组)、anti-miR-508-3p(转染miR-508-3p抑制剂, anti-miR-508-3p组)、anti-miR-NC(转染抑制剂阴性对照anti-miR-NC,anti-miR-NC组)、miR-508-3p+pcDNA-PAX2 (共转染miR-508-3p和PAX2过表达质粒pcDNA-PAX2, miR-508-3p+pcDNA-PAX2组)、miR-508-3p+pcDNA-NC(共转染miR-508-3p和空白质粒pcDNA-NC,miR-508-3p+pcDNA-NC组)。48 h后采用定量聚合酶链反应(qPCR)法或Western blotting检测转染后的细胞miR-508-3p或PAX2表达。
1.4 qPCR检测miR-508-3p、PAX2 mRNA表达
根据制造商的规程,使用TRIzol试剂提取组织或细胞总RNA。使用PrimeScript RT Reagent Kit合成互补DNA。使用SYBR Green qPCR Master Mix在ABI 7500系统上进行qPCR反应。使用的引物序列如下: miR-508-3p正向5′-CAAGCATGATTGTAGCCTTTTG-3′, 反向5′-TATCGTTGTACTCCAGACCAAGAC-3′; U6小核RNA(snRNA)正向5′-CTCGCTTCGGCAGCACA-3′, 反向5′-AACGCTTCACGAATTTGCGT-3′。PAX2正向5′-CCTGGCCACACCATTGTTC-3′, 反向5′-TCACGTTTCCTCTTCTCACCAT-3′; GAPDH正向5′-CTCAAGATCATCAGCAATGCC-3′, 反向5′-GGTCATGAGTCCTTCCACGATAC-3′。使用2-ΔΔCt方法,以U6 snRNA和GAPDH作为内部对照分析miR-508-3p、PAX2 mRNA相对表达水平。
1.5 Western blotting检测蛋白表达
采用蛋白提取试剂盒提取组织或细胞总蛋白,以BCA法定量蛋白,蛋白于100 ℃变性8 min, 每孔道上样等量蛋白,进行十二烷基磺酸钠-聚丙烯酰胺凝胶电泳(SDS-PAGE), 电泳结束后将蛋白转膜至PVDF膜,膜置于5%脱脂奶粉溶液中封闭,室温摇床上慢摇1.5 h, 根据预染marker将PVDF膜含PAX2、MMP-2、Bcl-2和Bax的部分依次剪下,放入一抗溶液中(稀释倍数1∶ 1 000), 4 ℃孵育过夜, TBST缓冲液洗膜,随后于二抗溶液中室温孵育45 min, 洗膜,电化学发光(ECL)显影,曝光拍照。用ImageJ软件定量强度,内参为GAPDH。实验重复3次。
1.6 MTT法检测细胞增殖活力
胰酶消化对数生长期A549细胞,制备成细胞悬液,并调整浓度为1×104个/mL, 按照5×103个/孔接种细胞于96孔板,每孔加100 μL, 每个样本设置5个平行孔,于培养48 h吸掉培养液,每孔中加10 μL MTT溶液,温育4 h, 中加150 μL DMSO, 平板摇床上摇匀后,采用酶标仪于490 nm处测定光密度(OD)值。
1.7 Transwell小室检测细胞侵袭能力
将Transwell小室置于24孔板内,向每孔中加100 μL已配置好的基质胶工作液,来回晃动培养板,及时将气泡去除,使基质胶均匀包被在小室的上室,于培养箱中常规培养1 h。用胰酶消化经si-PAX2处理48 h的A549细胞,制备成单细胞悬液,计数。小室上室中加300 μL细胞悬液(5×105个细胞),相应的每孔下室中加600 μL含FBS的完全培养基,然后放入培养箱中常规培养。24 h后,取出Transwell小室,使用棉签轻轻擦掉上室内细胞、液体及基质胶。用甲醇固定, 0.1%结晶紫染色, 15~20 min后,用PBS缓冲液将多余结晶紫洗去,风干。倒置显微镜下观察小室外底膜上染色后呈现的阳性细胞,拍照。显微镜下随机选择5个视野,取均值,实验重复3次。
1.8 流式细胞术检测细胞凋亡率
以1×105个/孔密度接种对数生长期A549细胞于6孔细胞培养板中, 24 h吸弃原有培养基,转染si-PAX2, 48 h后,用胰酶消化细胞,消化终止后将细胞转移至流式管,离心,收集细胞。加500 μL结合缓冲液重悬细胞,然后加5 μL碘化丙啶(PI), 混匀,再加入5 μL膜联蛋白V-异硫氰酸荧光素(Annexin V-FITC), 混匀,室温避光孵育15 min, 以流式细胞术仪检测分析。实验重复3次。
1.9 双荧光素酶活性检测
分别构建含有miR-508-3p结合位点的PAX2 -3′UTR-野生型(WT)及突变型(MUT)报告基因载体,在A549细胞中转染miR-508-3p和PAX2 -3′UTR-WT及MUT报告基因载体。48 h后收集细胞采用双荧光素酶报告基因检测系统检测荧光素酶活性。
1.10 统计学分析
本研究实验数据均采用SPSS 21.0软件进行分析,计量资料以(x±s)表示, 2组差异比较采用独立样本t检验,多组差异比较采用单因素方差分析,两两比较采用LSD-t检验,以P < 0.05为差异有统计学意义。
2. 结果
2.1 肺癌组织中miR-508-3p、PAX2 mRNA、PAX2蛋白表达水平和相关性分析
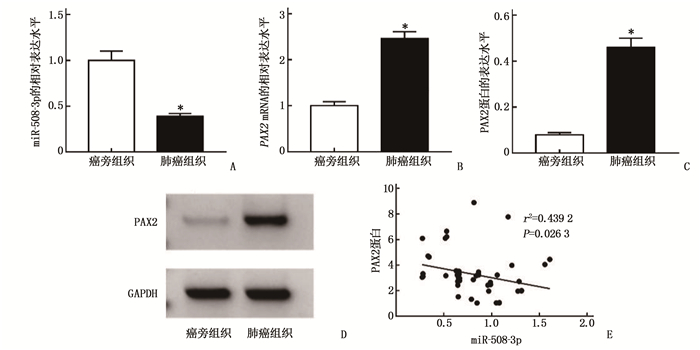
与癌旁组织比较,肺癌组织中miR-508-3p相对表达水平降低, PAX2 mRNA相对表达水平和PAX2蛋白表达水平升高,差异有统计学意义(P < 0.05); PAX2蛋白表达水平与miR-508-3p表达水平呈显著负相关(P < 0.05)。见图 1。
![]() 图 1 miR-508-3p、PAX2 mRNA、PAX2蛋白表达水平和相关性分析A: miR-508-3p相对表达水平柱状图; B: PAX2 mRNA相对表达水平柱状图; C: PAX2蛋白表达水平柱状图; D: PAX2蛋白Western blotting条带图; E: PAX2蛋白与miR-508-3p的相关性分析图。与癌旁组织比较, *P < 0.05。
图 1 miR-508-3p、PAX2 mRNA、PAX2蛋白表达水平和相关性分析A: miR-508-3p相对表达水平柱状图; B: PAX2 mRNA相对表达水平柱状图; C: PAX2蛋白表达水平柱状图; D: PAX2蛋白Western blotting条带图; E: PAX2蛋白与miR-508-3p的相关性分析图。与癌旁组织比较, *P < 0.05。2.2 转染miR-508-3p对肺癌A549细胞增殖活力、侵袭能力和细胞凋亡的影响
与空白组比较, miR-508-3p组miR-508-3p、Bax表达水平和细胞凋亡率均升高, MMP-2、Bcl-2蛋白表达水平和增殖活力(OD值)、侵袭能力均降低,差异有统计学意义(P < 0.05); miR-NC组与空白组比较,差异无统计学意义(P>0.05)。由此提示,转染miR-508-3p可抑制肺癌A549细胞增殖活力和侵袭能力,诱导细胞凋亡。见图 2、表 1。
![]() 图 2 转染miR-508-3p对肺癌A549细胞增殖活力、侵袭能力、细胞凋亡的影响A: 3组miR-508-3p相对表达水平(与空白组比较, *P < 0.05); B: 3组细胞凋亡情况的流式细胞仪检测结果; C: 3组细胞侵袭情况的Transwell检测结果; D: 3组MMP-2、Bcl-2和Bax蛋白Western blotting条带图。表 1 空白组、miR-NC组、miR-508-3p组蛋白表达、细胞增殖活力、侵袭能力和细胞凋亡结果比较(x±s)
图 2 转染miR-508-3p对肺癌A549细胞增殖活力、侵袭能力、细胞凋亡的影响A: 3组miR-508-3p相对表达水平(与空白组比较, *P < 0.05); B: 3组细胞凋亡情况的流式细胞仪检测结果; C: 3组细胞侵袭情况的Transwell检测结果; D: 3组MMP-2、Bcl-2和Bax蛋白Western blotting条带图。表 1 空白组、miR-NC组、miR-508-3p组蛋白表达、细胞增殖活力、侵袭能力和细胞凋亡结果比较(x±s)组别 n miR-508-3p MMP-2 Bcl-2 Bax OD值 侵袭能力 细胞凋亡率/% 空白组 9 1.00±0.07 0.69±0.07 0.33±0.03 0.08±0.01 0.98±0.07 168.35±6.31 4.23±0.82 miR-NC组 9 1.03±0.10 0.68±0.05 0.31±0.03 0.09±0.01 0.98±0.07 162.17±6.13 4.19±0.75 miR-508-3p组 9 2.16±0.13* 0.19±0.02* 0.12±0.01* 0.27±0.02* 0.54±0.04* 85.89±2.13* 27.02±1.83* MMP-2: 基质金属蛋白酶-2; Bcl-2: B淋巴细胞瘤-2; Bax: Bcl-2相关X蛋白; OD: 光密度。与空白组比较, * P < 0.05。 2.3 si-PAX2对肺癌A549细胞增殖活力、侵袭能力和细胞凋亡的影响
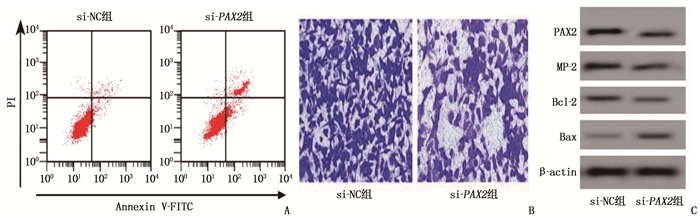
与si-NC组比较, si-PAX2组PAX2、MMP-2、Bcl-2表达水平降低, Bax表达水平、凋亡率升高,增殖活力和侵袭能力降低,差异有统计学意义(P < 0.05), 提示使用设计合成的PAX2特异性siRNA转染48 h, 可抑制肺癌A549细胞增殖活力和侵袭能力,诱导细胞凋亡。见图 3、表 2。
![]() 图 3 si-PAX2对肺癌A549细胞侵袭能力和蛋白表达的影响A: 2组细胞凋亡情况的流式细胞仪检测结果; B: 2组细胞侵袭情况的Transwell检测结果; C: 2组PAX2、MMP-2、Bcl-2和Bax蛋白Western blotting条带图。表 2 si-NC组、si-PAX2组蛋白表达、细胞增殖活力、侵袭能力和细胞凋亡结果比较(x±s)
图 3 si-PAX2对肺癌A549细胞侵袭能力和蛋白表达的影响A: 2组细胞凋亡情况的流式细胞仪检测结果; B: 2组细胞侵袭情况的Transwell检测结果; C: 2组PAX2、MMP-2、Bcl-2和Bax蛋白Western blotting条带图。表 2 si-NC组、si-PAX2组蛋白表达、细胞增殖活力、侵袭能力和细胞凋亡结果比较(x±s)组别 n PAX2 MMP-2 Bcl-2 Bax OD值 侵袭能力 细胞凋亡率/% si-NC组 9 0.72±0.07 0.67±0.07 0.32±0.03 0.07±0.01 0.97±0.07 166.45±5.72 4.21±0.18 si-PAX2组 9 0.20±0.02* 0.19±0.02* 0.20±0.02* 0.20±0.02* 0.66±0.06* 118.18±3.43* 20.13±1.26* PAX2 : 配对盒基因2; MMP-2: 基质金属蛋白酶-2; Bcl-2: B淋巴细胞瘤-2; Bax: Bcl-2相关X蛋白; OD: 光密度。与si-NC组比较, * P < 0.05。 2.4 miR-508-3p对PAX2的靶向调控作用
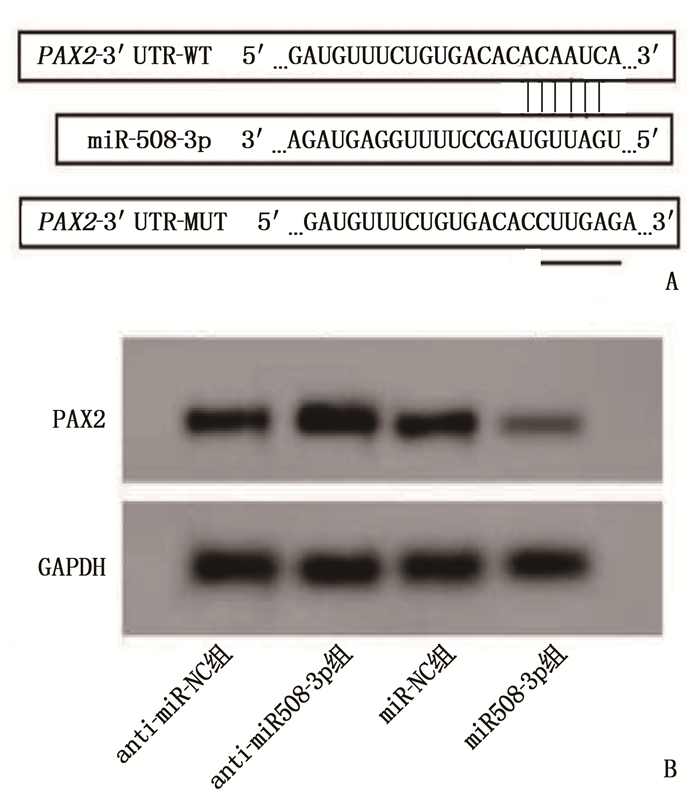
Starbase预测结果显示, miR-508-3p含有PAX2的互补序列,见图 4A。双荧光素酶活性检测和Western blotting检测结果显示,与miR-NC组比较, miR-508-3p组转染PAX2-3′UTR-WT的A549细胞荧光素酶活性降低, PAX2表达水平降低,差异有统计学意义(P < 0.05); miR-508-3p组转染PAX2 -3′UTR-MUT的A549细胞荧光素酶活性与miR-NC组比较,差异无统计学意义(P>0.05); anti-miR-508-3p组PAX2表达水平高于anti-miR-NC组,差异有统计学意义(P < 0.05)。见图 4B、表 3、表 4。由此提示, miR-508-3p可靶向PAX2基因,调控PAX2蛋白表达水平。
![]() 图 4 miR-508-3p靶向调控PAX2的预测图和PAX2蛋白表达水平A: PAX2与miR-508-3p结合的starbase预测图; B: 4组PAX2蛋白Western blotting条带图。表 3 miR-NC组、miR-508-3p组双荧光素酶活性检测结果(x±s)
图 4 miR-508-3p靶向调控PAX2的预测图和PAX2蛋白表达水平A: PAX2与miR-508-3p结合的starbase预测图; B: 4组PAX2蛋白Western blotting条带图。表 3 miR-NC组、miR-508-3p组双荧光素酶活性检测结果(x±s)组别 n 荧光素酶活性 WT MUT miR-NC组 9 1.00±0.10 1.00±0.08 miR-508-3p组 9 0.41±0.03* 0.97±0.07 WT: 野生型; MUT: 突变型。与miR-NC组比较, * P < 0.05。 表 4 4组PAX2蛋白表达水平比较(x±s)组别 n PAX2 anti-miR-NC组 9 0.71±0.07 anti-miR-508-3p组 9 1.24±0.10* miR-NC组 9 0.70±0.06 miR-508-3p组 9 0.29±0.03# PAX2 : 配对盒基因2。与anti-miR-NC组比较, * P < 0.05; 与miR-NC组比较, #P < 0.05。 2.5 PAX2逆转miR-508-3p对肺癌A549细胞增殖活力、侵袭能力、细胞凋亡的影响
与miR-508-3p+pcDNA-NC组比较, miR-508-3p+pcDNA-PAX2组PAX2、MMP-2、Bcl-2表达水平升高, Bax表达水平、细胞凋亡率降低,增殖活力和侵袭能力升高,差异均有统计学意义(P < 0.05), 表明PAX2可以逆转miR-508-3p对肺癌A549细胞增殖活力、侵袭能力及细胞凋亡的影响。见图 5、表 5。
![]() 图 5 2组细胞凋亡、侵袭能力和蛋白表达结果A: 2组细胞凋亡的流式细胞仪检测结果; B: 2组细胞侵袭数量的Transwell检测结果; C: 2组PAX2、MMP-2、Bcl-2和Bax蛋白Western blotting条带图。表 5 2组蛋白表达、细胞增殖活力、侵袭能力及细胞凋亡情况比较(x±s)
图 5 2组细胞凋亡、侵袭能力和蛋白表达结果A: 2组细胞凋亡的流式细胞仪检测结果; B: 2组细胞侵袭数量的Transwell检测结果; C: 2组PAX2、MMP-2、Bcl-2和Bax蛋白Western blotting条带图。表 5 2组蛋白表达、细胞增殖活力、侵袭能力及细胞凋亡情况比较(x±s)组别 PAX2 MMP-2 Bcl-2 Bax OD值 侵袭能力 细胞凋亡率/% miR-508-3p+pcDNA-NC组 0.28±0.02 0.18±0.01 0.10±0.01 0.26±0.01 0.54±0.03 82.96±3.10 26.93±1.25 miR-508-3p+pcDNA-PAX2组 0.68±0.05* 0.53±0.04* 0.36±0.02* 0.08±0.01* 0.97±0.08* 149.32±1.01* 7.25±0.32* PAX2 : 配对盒基因2; MMP-2: 基质金属蛋白酶-2; Bcl-2: B淋巴细胞瘤-2; Bax: Bcl-2相关X蛋白; OD: 光密度。与miR-508-3p+pcDNA-NC组比较, * P < 0.05。 3. 讨论
miR-508-3p是多种肿瘤的重要调节因子,研究[10]报道, miR-508-3p模拟物可抑制卵巢癌细胞增殖、迁移和侵袭。miR-508-3p在三阴性乳腺癌组织和细胞中表达显著降低,miR-508-3p过表达通过上调锌指E-盒结合同源异形盒-1(ZEB1)显著抑制三阴性乳腺癌细胞的侵袭能力[11]。本研究结果发现, miR-508-3p在肺癌中同样低表达,其过表达具有抑制肺癌细胞增殖、侵袭以及促进凋亡的作用,提示miR-508-3p作为肺癌的肿瘤抑制因子参与肺癌进展。
研究[12]显示, miRNA通过靶向靶mRNA发挥细胞功能。本研究发现,肺癌组织中PAX2蛋白表达与miR-508-3p呈显著负相关,且Starbase生物信息预测结果和双荧光素酶报告实验确定PAX2为miR-508-3p的下游靶基因,且miR-508-3p负调控PAX2蛋白表达。PAX2定位于人类10号染色体q24-25上,是一种重要的转录因子,在发育成熟组织中,异常表达的PAX2可呈现致癌作用[13-14]。近年来,相关研究[15-16]报道了PAX2在卵巢癌、急性髓性白血病等疾病中的表达水平,并发现PAX2在不同肿瘤中可能发挥不同作用。本研究收集43例患者的肺癌组织标本,采用Western blotting检测组织中PAX2表达,结果显示肺癌组织中PAX2表达水平显著高于癌旁组织,与既往研究[4]结论一致。张丽萍等[17]研究显示,用PAX2 siRNA转染子宫内膜癌HEC-1A细胞,可明显抑制癌细胞增殖,促进细胞凋亡。UEDA T等[18]研究显示, PAX2可通过对HGF的转录调控而增强前列腺癌细胞的侵袭能力。由此推测,抑制PAX2表达或可影响肺癌细胞生长。本研究用设计合成的PAX2特异性siRNA转染肺癌A549细胞,转染后的细胞PAX2表达水平显著降低,进一步行生物学特性研究发现, PAX2表达被抑制后的A549细胞增殖能力和侵袭能力均显著降低,凋亡率显著升高。本研究还发现, PAX2可以逆转miR-508-3p对A549细胞增殖活力、侵袭能力以及细胞凋亡的影响,提示miR-508-3p对肺癌细胞的抑制作用与下调PAX2表达有关。
基质金属蛋白酶(MMP)是一个大家族,几乎能降解细胞外基质中的各种蛋白成分,在肿瘤侵袭及转录过程中发挥着关键性作用[19-20]。MMP-2为MMP家族成员之一,是肿瘤侵袭转移的标志分子,多项研究[21-22]表明,抑制MMP-2表达可降低肺癌细胞侵袭和转移能力,但PAX2能否通过调节MMP-2影响肺癌细胞侵袭能力尚未明确。在细胞凋亡过程中, Bcl-2和Bax分别发挥抑凋亡作用和促凋亡作用,两者可形成同源二聚体(Bcl-2/Bcl-2, Bax/Bax)或异源二聚体(Bcl-2/Bax), 从而促进或抑制细胞凋亡, Bcl-2和Bax是细胞死亡的关键调节因素[23-24]。多项研究[25-26]表明,调节肺癌细胞Bcl-2和Bax表达可影响癌细胞凋亡。本研究用miR-508-3p模拟物或PAX2特异性siRNA转染肺癌A549细胞后,采用Western blotting检测MMP-2、Bcl-2和Bax表达,结果显示, MMP-2和Bcl-2表达降低, Bax表达升高。由此提示,过表达miR-508-3p或抑制PAX2表达可通过下调MMP-2、Bcl-2表达和上调Bax表达影响肺癌细胞侵袭能力和凋亡。
综上所述, miR-508-3p在肺癌组织中低表达, PAX2在肺癌组织中高表达,过表达miR-508-3p通过靶向抑制肺癌A549细胞PAX2表达而降低细胞增殖和侵袭能力,诱导细胞凋亡,提示miR-508-3p和PAX2可能是肺癌治疗的潜在靶标。
-
![]()
图 1 miR-508-3p、PAX2 mRNA、PAX2蛋白表达水平和相关性分析
A: miR-508-3p相对表达水平柱状图; B: PAX2 mRNA相对表达水平柱状图; C: PAX2蛋白表达水平柱状图; D: PAX2蛋白Western blotting条带图; E: PAX2蛋白与miR-508-3p的相关性分析图。与癌旁组织比较, *P < 0.05。
![]()
图 2 转染miR-508-3p对肺癌A549细胞增殖活力、侵袭能力、细胞凋亡的影响
A: 3组miR-508-3p相对表达水平(与空白组比较, *P < 0.05); B: 3组细胞凋亡情况的流式细胞仪检测结果; C: 3组细胞侵袭情况的Transwell检测结果; D: 3组MMP-2、Bcl-2和Bax蛋白Western blotting条带图。
![]()
图 3 si-PAX2对肺癌A549细胞侵袭能力和蛋白表达的影响
A: 2组细胞凋亡情况的流式细胞仪检测结果; B: 2组细胞侵袭情况的Transwell检测结果; C: 2组PAX2、MMP-2、Bcl-2和Bax蛋白Western blotting条带图。
![]()
图 4 miR-508-3p靶向调控PAX2的预测图和PAX2蛋白表达水平
A: PAX2与miR-508-3p结合的starbase预测图; B: 4组PAX2蛋白Western blotting条带图。
![]()
图 5 2组细胞凋亡、侵袭能力和蛋白表达结果
A: 2组细胞凋亡的流式细胞仪检测结果; B: 2组细胞侵袭数量的Transwell检测结果; C: 2组PAX2、MMP-2、Bcl-2和Bax蛋白Western blotting条带图。
表 1 空白组、miR-NC组、miR-508-3p组蛋白表达、细胞增殖活力、侵袭能力和细胞凋亡结果比较(x±s)
组别 n miR-508-3p MMP-2 Bcl-2 Bax OD值 侵袭能力 细胞凋亡率/% 空白组 9 1.00±0.07 0.69±0.07 0.33±0.03 0.08±0.01 0.98±0.07 168.35±6.31 4.23±0.82 miR-NC组 9 1.03±0.10 0.68±0.05 0.31±0.03 0.09±0.01 0.98±0.07 162.17±6.13 4.19±0.75 miR-508-3p组 9 2.16±0.13* 0.19±0.02* 0.12±0.01* 0.27±0.02* 0.54±0.04* 85.89±2.13* 27.02±1.83* MMP-2: 基质金属蛋白酶-2; Bcl-2: B淋巴细胞瘤-2; Bax: Bcl-2相关X蛋白; OD: 光密度。与空白组比较, * P < 0.05。  下载: 导出CSV
下载: 导出CSV
表 2 si-NC组、si-PAX2组蛋白表达、细胞增殖活力、侵袭能力和细胞凋亡结果比较(x±s)
组别 n PAX2 MMP-2 Bcl-2 Bax OD值 侵袭能力 细胞凋亡率/% si-NC组 9 0.72±0.07 0.67±0.07 0.32±0.03 0.07±0.01 0.97±0.07 166.45±5.72 4.21±0.18 si-PAX2组 9 0.20±0.02* 0.19±0.02* 0.20±0.02* 0.20±0.02* 0.66±0.06* 118.18±3.43* 20.13±1.26* PAX2 : 配对盒基因2; MMP-2: 基质金属蛋白酶-2; Bcl-2: B淋巴细胞瘤-2; Bax: Bcl-2相关X蛋白; OD: 光密度。与si-NC组比较, * P < 0.05。
下载: 导出CSV
表 3 miR-NC组、miR-508-3p组双荧光素酶活性检测结果(x±s)
组别 n 荧光素酶活性 WT MUT miR-NC组 9 1.00±0.10 1.00±0.08 miR-508-3p组 9 0.41±0.03* 0.97±0.07 WT: 野生型; MUT: 突变型。与miR-NC组比较, * P < 0.05。
下载: 导出CSV
表 4 4组PAX2蛋白表达水平比较(x±s)
组别 n PAX2 anti-miR-NC组 9 0.71±0.07 anti-miR-508-3p组 9 1.24±0.10* miR-NC组 9 0.70±0.06 miR-508-3p组 9 0.29±0.03# PAX2 : 配对盒基因2。与anti-miR-NC组比较, * P < 0.05; 与miR-NC组比较, #P < 0.05。
下载: 导出CSV
表 5 2组蛋白表达、细胞增殖活力、侵袭能力及细胞凋亡情况比较(x±s)
组别 PAX2 MMP-2 Bcl-2 Bax OD值 侵袭能力 细胞凋亡率/% miR-508-3p+pcDNA-NC组 0.28±0.02 0.18±0.01 0.10±0.01 0.26±0.01 0.54±0.03 82.96±3.10 26.93±1.25 miR-508-3p+pcDNA-PAX2组 0.68±0.05* 0.53±0.04* 0.36±0.02* 0.08±0.01* 0.97±0.08* 149.32±1.01* 7.25±0.32* PAX2 : 配对盒基因2; MMP-2: 基质金属蛋白酶-2; Bcl-2: B淋巴细胞瘤-2; Bax: Bcl-2相关X蛋白; OD: 光密度。与miR-508-3p+pcDNA-NC组比较, * P < 0.05。
下载: 导出CSV
-
[1] LI Q, CHEN K, DONG R, et al. LncRNA CASC2 inhibits autophagy and promotes apoptosis in non-small cell lung cancer cells via regulating the miR-214/TRIM16 axis[J]. RSC Adv, 2018, 8(71): 40846-40855. doi: 10.1039/C8RA09573F
[2] 刘小花, 肖春芳, 刘林秀. 血清NSE、CYFRA21-1、SCC、CEA联合检测在肺癌诊断中的临床应用[J]. 实用临床医药杂志, 2020, 24(19): 12-14. doi: 10.7619/jcmp.202019004 [3] 齐媛, 郭宝良. microRNA-361-5p在人类恶性肿瘤中的研究进展[J]. 实用临床医药杂志, 2021, 25(2): 117-121. doi: 10.7619/jcmp.20201614 [4] HUANG T, KANG W, ZHANG B, et al. miR-508-3p concordantly silences NFKB1 and RELA to inactivate canonical NF-κB signaling in gastric carcinogenesis[J]. Mol Cancer, 2016, 15: 9. doi: 10.1186/s12943-016-0493-7
[5] ZHAO L J, WANG W, XU L, et al. Integrative network biology analysis identifies miR-508-3p as the determinant for the mesenchymal identity and a strong prognostic biomarker of ovarian cancer[J]. Oncogene, 2019, 38(13): 2305-2319. doi: 10.1038/s41388-018-0577-5
[6] LIU P, GAO Y, HUAN J, et al. Upregulation of PAX2 promotes the metastasis of esophageal cancer through interleukin-5[J]. Cell Physiol Biochem, 2015, 35(2): 740-754. doi: 10.1159/000369734
[7] REN Y, HOU J, XU A, et al. Diagnostic utility of PAX2 and PAX5 in distinguishing non-small cell lung cancer from small cell lung cancer[J]. Int J Clin Exp Pathol, 2015, 8(11): 14709-14716. http://www.ijcep.com/files/ijcep0015449.pdf
[8] RUÍZ G, VALENCIA-GONZÁLEZ H A, LEÓN-GALICIA I, et al. Inhibition of RAD51 by siRNA and resveratrol sensitizes cancer stem cells derived from HeLa cell cultures to apoptosis[J]. Stem Cells Int, 2018, 2018: 2493869. http://downloads.hindawi.com/journals/sci/2018/2493869.pdf
[9] LU B, ZHOU Y, SU Z, et al. Effect of CCL2 siRNA on proliferation and apoptosis in the U251 human glioma cell line[J]. Mol Med Rep, 2017, 16(3): 3387-3394. doi: 10.3892/mmr.2017.6995
[10] GUO F, ZHANG K, LI M Y, et al. miR-508-3p suppresses the development of ovarian carcinoma by targeting CCNA2 and MMP7[J]. Int J Oncol, 2020, 57(1): 264-276. doi: 10.3892/ijo.2020.5055
[11] GUO S J, ZENG H X, HUANG P, et al. miR-508-3p inhibits cell invasion and epithelial-mesenchymal transition by targeting ZEB1 in triple-negative breast cancer[J]. Eur Rev Med Pharmacol Sci, 2018, 22(19): 6379-6385. http://www.europeanreview.org/wp/wp-content/uploads/6379-6385.pdf
[12] POWRÓZEK T, MA?ECKA-MASSALSKA T. MiRNA and lung cancer radiosensitivity: a mini-review[J]. Eur Rev Med Pharmacol Sci, 2019, 23(19): 8422-8428. http://www.ncbi.nlm.nih.gov/pubmed/31646572
[13] UEDA T, ITO S, SHIRAISHI T, et al. Hyper-expression of PAX2 in human metastatic prostate tumors and its role as a cancer promoter in an in vitro invasion model[J]. Prostate, 2013, 73(13): 1403-1412. doi: 10.1002/pros.22687
[14] PUSHCHINA E V, VARAKSIN A A, OBUKHOV D K. The PAX2 and Pax6 transcription factors in the optic nerve and brain of trout oncorhynchus mykiss after a mechanical eye injury[J]. Russ J Dev Biol, 2018, 49(5): 264-290. doi: 10.1134/S1062360418050041
[15] SONG H, KWAN S Y, IZAGUIRRE D I, et al. PAX2 expression in ovarian cancer[J]. Int J Mol Sci, 2013, 14(3): 6090-6105. doi: 10.3390/ijms14036090
[16] SIEHL J M, THIEL E, HEUFELDER K, et al. Possible regulation of Wilms' tumour gene 1 (WT1) expression by the paired box genes PAX2 and PAX8 and by the haematopoietic transcription factor GATA-1 in human acute myeloid leukaemias[J]. Br J Haematol, 2015, 123(2): 235-242. http://www.researchgate.net/profile/Eckhard_Thiel2/publication/9060831_Possible_regulation_of_Wilms'_tumour_gene_1_WT1_expression_by_the_paired_box_genes_PAX2_and_PAX8_and_by_the_haematopoietic_transcription_factor_GATA-1_in_human_acute_myeloid_leukaemias/links/555b2b5108aeaaff3bfc1133.pdf
[17] 张丽萍, 石小燕, 刘永珍, 等. PAX2特异性siRNA在诱导HEC-1A细胞增殖及凋亡中的作用[J]. 华中科技大学学报: 医学版, 2012, 41(1): 84-87. doi: 10.3870/j.issn.1672-0741.2012.01.019 [18] UEDA T, ITO S, SHIRAISHI T, et al. PAX2 promoted prostate cancer cell invasion through transcriptional regulation of HGF in an in vitro model[J]. Biochim Biophys Acta, 2015, 1852(11): 2467-2473. doi: 10.1016/j.bbadis.2015.08.008
[19] BAI X Y, LI S, WANG M, et al. Krüppel-like factor 9 down-regulates matrix metalloproteinase 9 transcription and suppresses human breast cancer invasion[J]. Cancer Lett, 2018, 412: 224-235. doi: 10.1016/j.canlet.2017.10.027
[20] KHOJASTEHFARD M, DOLATKHAH H, SOMI M H, et al. The effect of oral administration of PUFAs on the matrix metalloproteinase expression in gastric adenocarcinoma patients undergoing chemotherapy[J]. Nutr Cancer, 2019, 71(3): 444-451. doi: 10.1080/01635581.2018.1506494
[21] CAO C, XU N, ZHENG X, et al. Elevated expression of MMP-2 and TIMP-2 cooperatively correlates with risk of lung cancer[J]. Oncotarget, 2017, 8(46): 80560-80567. doi: 10.18632/oncotarget.20156
[22] XUE M Q, SHI D F, XU G H, et al. The long noncoding RNA linc00858 promotes progress of lung cancer through miR-3182/MMP2 axis[J]. Artif Cells Nanomed Biotechnol, 2019, 47(1): 2091-2097. doi: 10.1080/21691401.2019.1617728
[23] 梁岚, 张洁. 甘草甜素对人食管癌细胞ECA109的影响及机制[J]. 中国老年学杂志, 2018, 38(4): 914-917. doi: 10.3969/j.issn.1005-9202.2018.04.060 [24] AL-QATHAMA A, GIBBONS S, PRIETO J M. Differential modulation of Bax/Bcl-2 ratio and onset of caspase-3/7 activation induced by derivatives of Justicidin B in human melanoma cells A375[J]. Oncotarget, 2017, 8(56): 95999-96012. doi: 10.18632/oncotarget.21625
[25] YUE H C, HUANG M, CHEN W, et al. N-acetylglucosaminyltransferase V modulates radiosensitivity and migration of small cell lung cancer through epithelial-mesenchymal transition[J]. Febs J, 2016, 282(22): 4295-4306. http://www.researchgate.net/profile/Miaojuan_Huang/publication/281169324_N-acetylglucosaminyltransferase_V_modulates_radiosensitivity_and_migration_of_small_cell_lung_cancer_through_epithelial-mesenchymal_transition/links/5789dc4608ae7a588ee92ff3.pdf
[26] LI CH, WU X, SUN R, et al. Croton tiglium extract induces apoptosis via bax/bcl-2 pathways in human lung cancer A549 cells[J]. Asian Pac J Cancer Prev, 2016, 17(11): 4893-4898. http://journal.waocp.org/article_41305_441366657a07afa94b9aa61ca8b5c70e.pdf
-
期刊类型引用(3)
1. 鲍祯,乔亚红. 纤维蛋白胶凝素-3在非小细胞肺癌中的表达及其生物学功能研究. 实用临床医药杂志. 2024(14): 18-23 .  本站查看
本站查看
2. 王刚,朱洪宽,陈永伦,李建钢. 丙泊酚调控miRNA-383-5p/CIP2A的癌性抑制因子轴抑制膀胱癌细胞恶性进展. 中国实验诊断学. 2023(03): 341-345 . 百度学术
3. 刘渊源,丁成智,吴晓明. 老年肺癌患者术后并发肺部感染的病原菌分布与耐药性分析. 抗感染药学. 2022(07): 963-967 . 百度学术
其他类型引用(0)


计量
- 文章访问数: 236
- HTML全文浏览量: 95
- PDF下载量: 10
- 被引次数: 3
 苏公网安备 32100302010246号
苏公网安备 32100302010246号